It looks like you're using an Ad Blocker.
Please white-list or disable AboveTopSecret.com in your ad-blocking tool.
Thank you.
Some features of ATS will be disabled while you continue to use an ad-blocker.

Ebola - my visual charts & projections based on WHO data
page: 5share:
WHO released new data today, covering the Ebola outbreak numbers through August 11, 2014. I've updated some of the charts I've been creating
below.
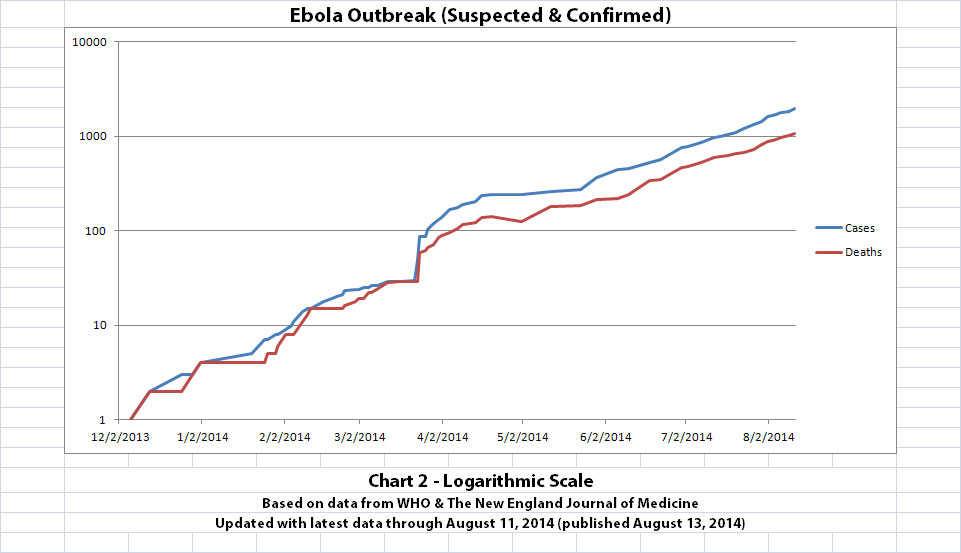
As with the original charts, Chart 1 and Chart 2 show the number of cases and the number of deaths for the Ebola outbreak that have actually been reported. In Chart 1, the y-axis is linear. In Chart 2, the y-axis is a logarithmic scale where divisions of the axis increase by powers of 10.
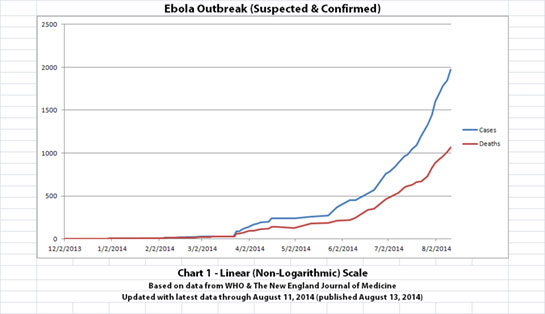
Click the graphic above to see it larger.
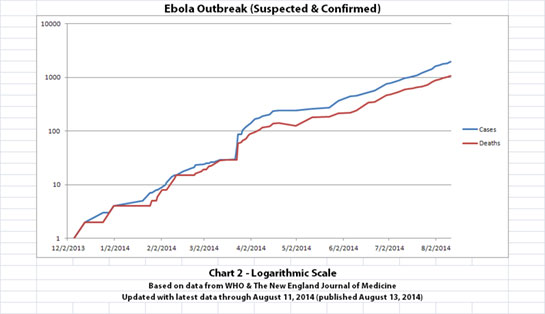
Click the graphic above to see it larger.
I didn't update charts 3, 4, and 5 (the projections) for two reasons. First, there wouldn't be much change yet, so previous projections in this thread are on-track, unfortunately. But second, I hope it's a good sign that all the suspected cases outside the original outbreak countries have allegedly been negative so far.
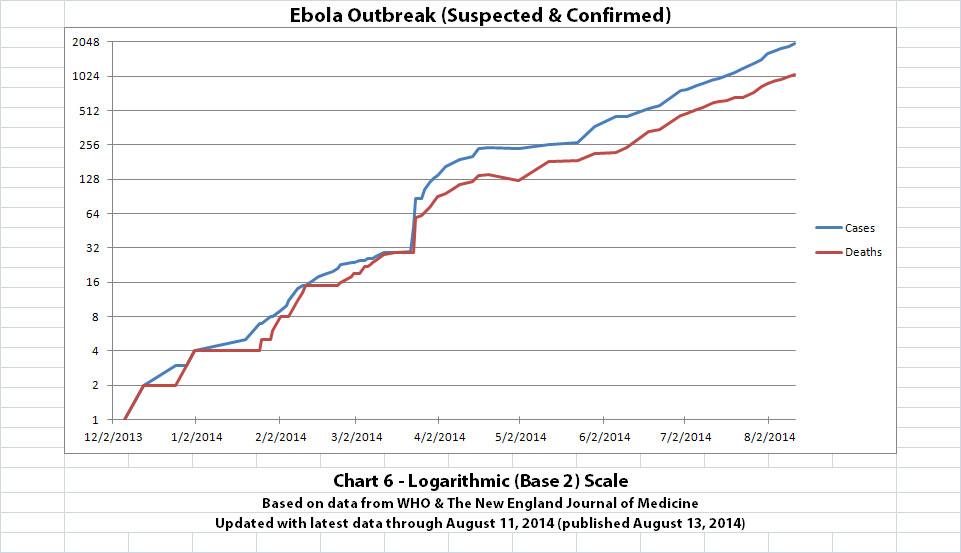
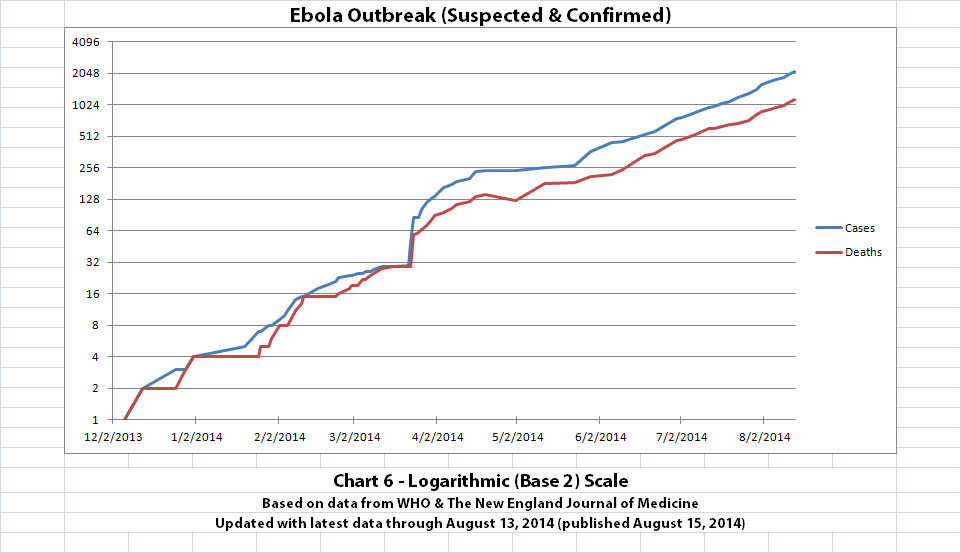
I updated the newest graph, Chart 6, with the new WHO data though. Chart 6 is similar to Chart 2 except that on Chart 6 the y-axis is in powers of 2 rather than powers of 10.
A number of people have mentioned or asked about the Basic Reproduction Number (R0), which is the average number of people who are infected by someone. I am not an epidemiologist. But if R0 is approximately 2 (give or take a little, as calculated from previous Ebola outbreaks) and if it is an average of about 2 or 3 weeks between generations (given the incubation time), I think Chart 6 shows about what one would expect. The current outbreak has doubled about 11 times in about 8 months and 11 days since the presumed first case.
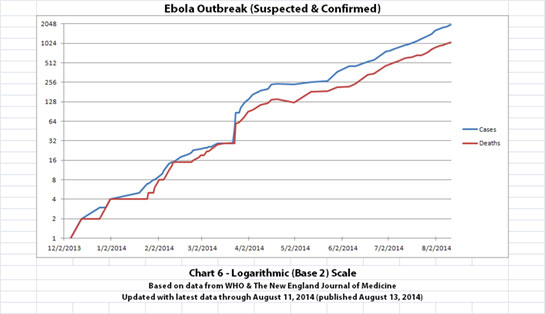
Click the graphic above to see it larger.
The charts above were created and updated by me (not by WHO or NEJM) using the data from the news updates on the following WHO and The New England Journal of Medicine websites:
SOURCE: WHO website 1
SOURCE: WHO website 2
SOURCE: The New England Journal of Medicine
As with the original charts, Chart 1 and Chart 2 show the number of cases and the number of deaths for the Ebola outbreak that have actually been reported. In Chart 1, the y-axis is linear. In Chart 2, the y-axis is a logarithmic scale where divisions of the axis increase by powers of 10.
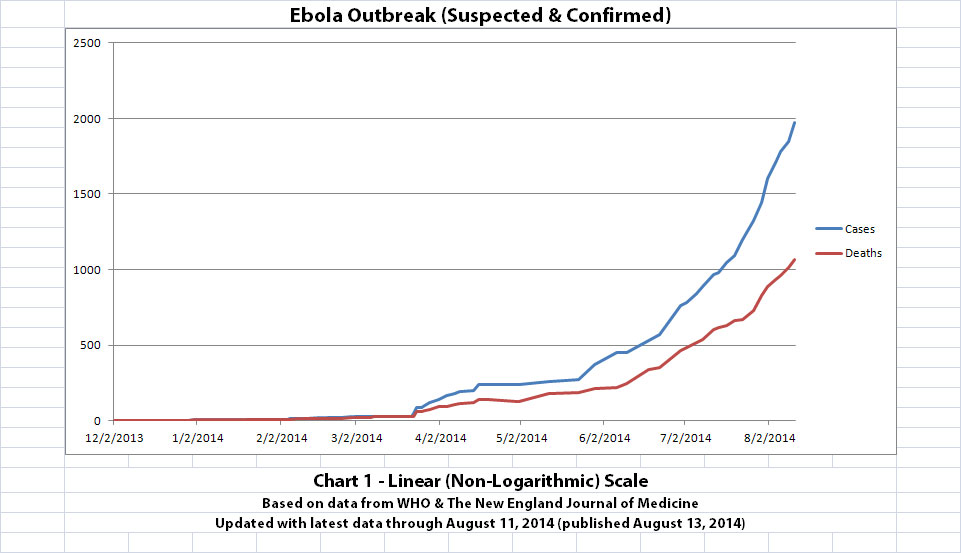
Click the graphic above to see it larger.
Click the graphic above to see it larger.
I didn't update charts 3, 4, and 5 (the projections) for two reasons. First, there wouldn't be much change yet, so previous projections in this thread are on-track, unfortunately. But second, I hope it's a good sign that all the suspected cases outside the original outbreak countries have allegedly been negative so far.
I updated the newest graph, Chart 6, with the new WHO data though. Chart 6 is similar to Chart 2 except that on Chart 6 the y-axis is in powers of 2 rather than powers of 10.
A number of people have mentioned or asked about the Basic Reproduction Number (R0), which is the average number of people who are infected by someone. I am not an epidemiologist. But if R0 is approximately 2 (give or take a little, as calculated from previous Ebola outbreaks) and if it is an average of about 2 or 3 weeks between generations (given the incubation time), I think Chart 6 shows about what one would expect. The current outbreak has doubled about 11 times in about 8 months and 11 days since the presumed first case.
Click the graphic above to see it larger.
The charts above were created and updated by me (not by WHO or NEJM) using the data from the news updates on the following WHO and The New England Journal of Medicine websites:
SOURCE: WHO website 1
SOURCE: WHO website 2
SOURCE: The New England Journal of Medicine
edit on
13-8-2014 by ikonoklast because: Changed Basic Reproduction Rate to Basic Reproduction Number.
ikonoclast you are doing a great job making these charts.
how come the scientific community did not do that in articles.
here are a collection of articles about ebola showing that
1- antibodies helped monkeys survive the disease.
2-that since 2003 outbreaks were composed of different strains at the same time?
no reason is found yet???
ebola collection of studies in science magazine. freecollection of all ebola science mag articles free
show antibodies saved lives of monkeys
and can do same in humans !
vaccine takes 6 months and could be dangerous and they been used in africa in trials. could the trials of vaccines caused the new outbreaks??
how come the scientific community did not do that in articles.
here are a collection of articles about ebola showing that
1- antibodies helped monkeys survive the disease.
2-that since 2003 outbreaks were composed of different strains at the same time?
no reason is found yet???
ebola collection of studies in science magazine. freecollection of all ebola science mag articles free
show antibodies saved lives of monkeys
and can do same in humans !
vaccine takes 6 months and could be dangerous and they been used in africa in trials. could the trials of vaccines caused the new outbreaks??
Epidemiology. Ebola outbreaks may have had independent sources.
ebola outbreak had multiple sources and strains in the past
also her, how is it possible one gorilla caracas have multiple strains at the same time. ebola so lethal to monkeys cant have several diseases at the same time or what is such probability??
one gorilla have multiple strains caused a previous outbreakThe human outbreaks consisted of multiple simultaneous epidemics caused by different viral strains, and each epidemic resulted from the handling of a distinct gorilla, chimpanzee, or duiker carcass
ebola outbreak had multiple sources and strains in the past
also her, how is it possible one gorilla caracas have multiple strains at the same time. ebola so lethal to monkeys cant have several diseases at the same time or what is such probability??
one gorilla have multiple strains caused a previous outbreakThe human outbreaks consisted of multiple simultaneous epidemics caused by different viral strains, and each epidemic resulted from the handling of a distinct gorilla, chimpanzee, or duiker carcass
edit on 13-8-2014 by
viibird because: (no reason given)
a reply to: ikonoklast
Ironiklast... Great charts!!!!
However until the cases/deaths ratio minimizes or becomes closes together... and the cases slows down and stops or decreases... i won't feel good about it.
Ironiklast... Great charts!!!!
However until the cases/deaths ratio minimizes or becomes closes together... and the cases slows down and stops or decreases... i won't feel good about it.
if the villages of source were quarantined along with their infected, the epidemic would have died on its own.
hospitals were the culprit in spreading the disease a fact known to cdc and others few years ago. why did they not i mplement the quarantine few months ago?
the 2010 study :Trends and Challenges in International Security:
An Inventory by Fred Schreier
2010 study pandemics logistics
quote "The crucial element in any response to a pandemic may
be the political will to impose quarantine, and to restrict travel and trade."
hospitals were the culprit in spreading the disease a fact known to cdc and others few years ago. why did they not i mplement the quarantine few months ago?
the 2010 study :Trends and Challenges in International Security:
An Inventory by Fred Schreier
2010 study pandemics logistics
quote "The crucial element in any response to a pandemic may
be the political will to impose quarantine, and to restrict travel and trade."
edit on 13-8-2014 by viibird because: (no reason given)
a reply to: viibird
I'm glad the charts are helpful, thanks for letting me know. Some of the links in your last couple posts in this thread are broken. Maybe you can edit those posts if you want to fix them.
Apparently having more than one strain of a virus in one body (human or animal) does happen and can lead to recombination, altering the gene sequence of a virus. This is supposedly what formed one of those nasty strains of flu a few years ago. It seems statistically unlikely two different strains of a rare and lethal virus like Ebola would be in one body at the same time, but apparently it is known to have happened:
Isolates of Zaire ebolavirus from wild apes reveal genetic lineage and recombinants
A deeper question might be how that could happen. I'm not an expert, but I can only think of two possible ways: naturally or artificially. Naturally seems statistically unlikely, but it's a strange world. As for artificially, I've read that this is one way new strains of viruses are created for biological weapons. soficrow has an ATS thread about there being bio-warfare laboratories near the current original outbreak of this Ebola and the outbreak in Nigeria.
I think the outbreak in Nigeria probably traces to that guy, Sawyer. But as for the original outbreak this time, we'll probably never know.
I'm glad the charts are helpful, thanks for letting me know. Some of the links in your last couple posts in this thread are broken. Maybe you can edit those posts if you want to fix them.
Apparently having more than one strain of a virus in one body (human or animal) does happen and can lead to recombination, altering the gene sequence of a virus. This is supposedly what formed one of those nasty strains of flu a few years ago. It seems statistically unlikely two different strains of a rare and lethal virus like Ebola would be in one body at the same time, but apparently it is known to have happened:
Isolates of Zaire ebolavirus from wild apes reveal genetic lineage and recombinants
A deeper question might be how that could happen. I'm not an expert, but I can only think of two possible ways: naturally or artificially. Naturally seems statistically unlikely, but it's a strange world. As for artificially, I've read that this is one way new strains of viruses are created for biological weapons. soficrow has an ATS thread about there being bio-warfare laboratories near the current original outbreak of this Ebola and the outbreak in Nigeria.
I think the outbreak in Nigeria probably traces to that guy, Sawyer. But as for the original outbreak this time, we'll probably never know.
so artificially most likely considering one of dr doom hardline believers injected or did spread the different strains in different persons. [Dr doom
pianka in 2006 promote ebola to cut humans to 10%!!!???dr doom
eric pianka a disabled man promote ebola in 2006 to cut population to
10%www.evolutionnews.org...
[/url]http://www.evolutionnews.org/2006/04/doctor_doom_eric_pianka_receiv002118.html
dr doom dr eric pianka advocated the elimination of 90 percent of Earth's population by airborne Ebola in 2006.!!!
he was even investigated by FBI then.
that will show the curves overlapping each other and showing as one huge tsunami ever rising curve.
if it was a one strain then there will be curves separated by time just like econoklast presented of the old study of 1995 2000 outbreaks.
how can people eat a roated caracas of a gorilla. the gorilla must have died of one strain of one incubation period( like 1995,2000 )
the study by Schreier 2010 about Pandemics future and political will to quarantine, seems now is the time to do quarantine dspace.africaportal.org...
as for the collection of ebola studies showing antibodies effectiveness ebola studies collection at science mag www.sciencemag.org...
multiple strains in one gorilla???
www.ncbi.nlm.nih.gov...
dr doom dr eric pianka advocated the elimination of 90 percent of Earth's population by airborne Ebola in 2006.!!!
he was even investigated by FBI then.
that will show the curves overlapping each other and showing as one huge tsunami ever rising curve.
if it was a one strain then there will be curves separated by time just like econoklast presented of the old study of 1995 2000 outbreaks.
how can people eat a roated caracas of a gorilla. the gorilla must have died of one strain of one incubation period( like 1995,2000 )
edit on
13-8-2014 by viibird because: (no reason given)
the study by Schreier 2010 about Pandemics future and political will to quarantine, seems now is the time to do quarantine dspace.africaportal.org...
edit on 13-8-2014 by viibird because: (no reason given)
edit on
13-8-2014 by viibird because: (no reason given)
as for the collection of ebola studies showing antibodies effectiveness ebola studies collection at science mag www.sciencemag.org...
edit on 13-8-2014 by viibird because: (no reason given)
multiple strains in one gorilla???
www.ncbi.nlm.nih.gov...
edit on 13-8-2014 by viibird because: (no reason given)
Every thing about Ebola from a note to Doctors
Ebola virus incubates in infected humans for 2-21 days, with the majority of patients becoming symptomatic after 8-9 days. Once infected, patients can experience severe symptoms within 1-2 days.
Symptoms of Ebola include:
• Sudden fever, often as high as 103º-105º F;
• Intense weakness, sore throat, and headache; and
• Profuse vomiting and diarrhea (occurs 1-2 days after the aforementioned symptoms).
More severe symptoms, such as the development of coagulopathy with thrombocytopenia, can develop in as soon as 24-48 hours, leading to bleeding from the nasal or oral cavities, along with hemorrhagic skin blisters. The development of renal failure, leading to multisystem organ failure along with disseminated intravascular coagulation, can then rapidly ensue over 3-5 days, along with significant volume loss.
Patients who develop a fulminant course often die within 8-9 days. Those who survive beyond 2 weeks have a better prognosis for survival.
The Difficulties of Ebola Diagnoses
One of the difficulties encountered in identifying Ebola virus is that in the early days of the disease, the symptoms may be similar to those of other types of infectious diseases, such as malaria, Lassa fever, typhoid, cholera, and even meningitis. Only after 3-5 days (or even later in the course of the disease) might the hemorrhagic blisters -- along with internal hemorrhage, the hallmark of the illness -- become evident.
Ebola, because it is not airborne or spread by droplets, is not nearly as contagious as measles or influenza. Patients with measles or influenza can spread the virus before they are symptomatic, as opposed to those with Ebola, who are not infectious until symptoms have developed. Furthermore, direct contact with infected secretions, such as saliva, is essential to transmitting the Ebola virus. You cannot acquire Ebola virus if another person coughs or sneezes close to you, and it is not spread by casual contact. Rather, it is acquired by direct contact with infected secretions such as vomit, diarrhea, and blood primarily. It may also be spread by direct contact with saliva, sweat, and tears. Other means of transmission include contact of secretions with a skin opening or healing wound, or if a person contacts secretions and touches his or her eyes, nose, or mouth.
It is important to remember that only patients who are symptomatic are contagious and can then transmit the virus to others through their secretions. Those who have contracted the disease are primarily healthcare workers caring for patients, as well as family members who have had close contact with infected patients. Another method of infection has involved family members who handle corpses at the time of burial, along with those who eat fruit bats, antelope, or other animals potentially infected with the virus.
Studies indicate that the virus is in much higher concentration in vomit, blood, and diarrhea compared with saliva, sweat, and tears, making disinfection of public areas such as restrooms imperative in order to contain the virus.
The actual risk to citizens living and working in the United States is quite low
Treatment-Therapy
At this time, only supportive care is available (intravenous fluids; blood and platelet transfusions), although upcoming human vaccine trials may be promising.
The National Institutes of Health will begin a human vaccine trial in September 2014, according to recent statements from Dr. Anthony Fauci, Director of the National Institute of Allergy and Infectious Diseases (NIAID). Previous attempts at a human vaccine in the early 2000s were not successful.[3]
The current vaccine, developed by the NIAID Vaccine Research Center, contains no infectious Ebola virus material. It is actually a chimpanzee adenovirus vector vaccine that has incorporated 2 Ebola virus genes. Adenovirus vectors are useful delivery models as vaccines because the virus can be easily manipulated. As a nonreplicating viral vector, the vaccine works by entering a cell and delivering the new genetic material. The new genes that are inserted cause a protein to become expressed, which in turn produces an immune response in the body. According to NIAID, the vaccine has shown early promise in a primate model.
Another approach to help infected patients involves transfusing blood or plasma from those patients who have recently recovered from Ebola virus infection. This approach is based on the premise that the plasma from recovered patients contains life-saving antibodies. This is an experimental treatment that has been used, according to recent reports during this epidemic, although results of such treatment have not been formally reported.
Use of an experimental compound, referred to as BCX4430, was reported in the journal Nature in April 2014.[4] The compound, an RNA-dependent RNA polymerase inhibitor, has proven successful in a nonhuman primate model, whereby postexposure prophylaxis to BCX4430 prevented death in 17 of 18 macaques studied. No human trials have yet been reported.
www.medscape.com...
Ebola virus incubates in infected humans for 2-21 days, with the majority of patients becoming symptomatic after 8-9 days. Once infected, patients can experience severe symptoms within 1-2 days.
Symptoms of Ebola include:
• Sudden fever, often as high as 103º-105º F;
• Intense weakness, sore throat, and headache; and
• Profuse vomiting and diarrhea (occurs 1-2 days after the aforementioned symptoms).
More severe symptoms, such as the development of coagulopathy with thrombocytopenia, can develop in as soon as 24-48 hours, leading to bleeding from the nasal or oral cavities, along with hemorrhagic skin blisters. The development of renal failure, leading to multisystem organ failure along with disseminated intravascular coagulation, can then rapidly ensue over 3-5 days, along with significant volume loss.
Patients who develop a fulminant course often die within 8-9 days. Those who survive beyond 2 weeks have a better prognosis for survival.
The Difficulties of Ebola Diagnoses
One of the difficulties encountered in identifying Ebola virus is that in the early days of the disease, the symptoms may be similar to those of other types of infectious diseases, such as malaria, Lassa fever, typhoid, cholera, and even meningitis. Only after 3-5 days (or even later in the course of the disease) might the hemorrhagic blisters -- along with internal hemorrhage, the hallmark of the illness -- become evident.
Ebola, because it is not airborne or spread by droplets, is not nearly as contagious as measles or influenza. Patients with measles or influenza can spread the virus before they are symptomatic, as opposed to those with Ebola, who are not infectious until symptoms have developed. Furthermore, direct contact with infected secretions, such as saliva, is essential to transmitting the Ebola virus. You cannot acquire Ebola virus if another person coughs or sneezes close to you, and it is not spread by casual contact. Rather, it is acquired by direct contact with infected secretions such as vomit, diarrhea, and blood primarily. It may also be spread by direct contact with saliva, sweat, and tears. Other means of transmission include contact of secretions with a skin opening or healing wound, or if a person contacts secretions and touches his or her eyes, nose, or mouth.
It is important to remember that only patients who are symptomatic are contagious and can then transmit the virus to others through their secretions. Those who have contracted the disease are primarily healthcare workers caring for patients, as well as family members who have had close contact with infected patients. Another method of infection has involved family members who handle corpses at the time of burial, along with those who eat fruit bats, antelope, or other animals potentially infected with the virus.
Studies indicate that the virus is in much higher concentration in vomit, blood, and diarrhea compared with saliva, sweat, and tears, making disinfection of public areas such as restrooms imperative in order to contain the virus.
The actual risk to citizens living and working in the United States is quite low
Treatment-Therapy
At this time, only supportive care is available (intravenous fluids; blood and platelet transfusions), although upcoming human vaccine trials may be promising.
The National Institutes of Health will begin a human vaccine trial in September 2014, according to recent statements from Dr. Anthony Fauci, Director of the National Institute of Allergy and Infectious Diseases (NIAID). Previous attempts at a human vaccine in the early 2000s were not successful.[3]
The current vaccine, developed by the NIAID Vaccine Research Center, contains no infectious Ebola virus material. It is actually a chimpanzee adenovirus vector vaccine that has incorporated 2 Ebola virus genes. Adenovirus vectors are useful delivery models as vaccines because the virus can be easily manipulated. As a nonreplicating viral vector, the vaccine works by entering a cell and delivering the new genetic material. The new genes that are inserted cause a protein to become expressed, which in turn produces an immune response in the body. According to NIAID, the vaccine has shown early promise in a primate model.
Another approach to help infected patients involves transfusing blood or plasma from those patients who have recently recovered from Ebola virus infection. This approach is based on the premise that the plasma from recovered patients contains life-saving antibodies. This is an experimental treatment that has been used, according to recent reports during this epidemic, although results of such treatment have not been formally reported.
Use of an experimental compound, referred to as BCX4430, was reported in the journal Nature in April 2014.[4] The compound, an RNA-dependent RNA polymerase inhibitor, has proven successful in a nonhuman primate model, whereby postexposure prophylaxis to BCX4430 prevented death in 17 of 18 macaques studied. No human trials have yet been reported.
www.medscape.com...
Hey ikonoklast, thank you for the charts. I'm not good at math, and even I can see where this is going.
Also, a heads-up: WHO now says the reported cases & deaths are vastly underestimated - www.abovetopsecret.com...
Also, a heads-up: WHO now says the reported cases & deaths are vastly underestimated - www.abovetopsecret.com...
Ebola, because it is not airborne or spread by droplets, is not nearly as contagious as measles or influenza. Patients with measles or influenza can spread the virus before they are symptomatic, as opposed to those with Ebola, who are not infectious until symptoms have developed. Furthermore, direct contact with infected secretions, such as saliva, is essential to transmitting the Ebola virus. You cannot acquire Ebola virus if another person coughs or sneezes close to you, and it is not spread by casual contact. Rather, it is acquired by direct contact with infected secretions such as vomit, diarrhea, and blood primarily. It may also be spread by direct contact with saliva, sweat, and tears. Other means of transmission include contact of secretions with a skin opening or healing wound, or if a person contacts secretions and touches his or her eyes, nose, or mouth.
It is important to remember that only patients who are symptomatic are contagious and can then transmit the virus to others through their secretions.
Okay, I have major league issues with this. Are they really saying that if someone with an active case of Ebola, but not yet visually sick (they may feel like they're coming down with something but be too bewildered, ill and tired to comment upon it...) could sneeze on you, cough on you, and then you could touch your eyes with that mist of saliva on you and NOT get it?
At what point does 'not sick yet, not contagious' cross over into 'spreading around a lethal disease'? Is it safe at noon, deadly at 12:05 or what??
And are you willing to bet your life on the CDC and WHO's description of a virus that may well have been weaponized, manipulated, or undergone mutation itself?
Once someone is exposed, the virus is rapidly replicating inside them; I dunno about you, but if Biocontainment Level IV, the highest level complete with space suits and a room equipped with air pressure valves, where everything that exits is burned or otherwise decontaminated with a vengeance, is necessary to work with it, then I really don't think sitting next to someone incubating it on a plane for an overnight flight would be in the realm of a great idea...
And as I have stated in other posts, there isn't a hospital in the US or any Western country that is ready for this; they will be completely overwhelmed both staff-wise and supplies-wise in three, count 'em, THREE DAYS. Once the panic both gossiping and real sets in, the emergency rooms will be beset by hypochondriacs of all kinds, real flus and other diseases that look exactly the same at first will arrive in droves, and even the most altruistic staff will need to take a break after 36 hours or so. No guarantees at that point that replacements will arrive to work. There's precious little that Western medicine can do for those taken ill in any event, aside from fluid support. And we already have a shortage of that.
This is the outbreak the CDC has been having nightmares about for decades now; they had thought it was going to be H5N1, not unlike the Spanish Flu in its possible lethality. That it is Ebola is a thousand times worse. No one in the West has ever had this before, there are utterly no immunities, it isn't even known if having survived it once, you're immune afterwards (and you'll certainly be a pariah sexually for the rest of your life if you do) and we don't even know what the long term effects of survival may be; if the human body is permanently damaged by it or what. It would seem like it would cause such immune and organ issues that permanent disability would be a given. How can the kidneys or liver survive intact after an almost complete meltdown of the vascular system with a free-for-all of bleeding?
The 1918- 1919 Spanish Flu, the last bout we had with something like this, killed an estimated 20 to 50 million people in the West; numbers for Russia, China and South America were impossible to tally, people were too busy burying their dead and dying themselves to keep track. It killed more people in two years than died in all the wars of the 20th Century and killed more people in two years than died in three centuries of the Black Plague. The reason you've barely heard about it? Because the generation that lived through it didn't want to talk about it, they were that traumatized.
And the lethality of the Spanish Flu is about comparative with Ebola, as we know it. This may be a different entity altogether. If it's been fiddled with to slow down the incubation time, it will kill far more people because it will spread wider.
The old Ebola, the one no one in the West really worried about, never got out of the jungles, it couldn't walk out in time to infect new persons and was limited to patient zero who probably ate a bat or an infected monkey plus his cohorts. Even if it horrifically killed a whole village, the news barely made it out of the darkness that is the Congo.
This Ebola has planed, trained and bussed out of the jungle and is already global. Unless it self-mutates into something less lethal, always a possibility, we've got a long road ahead of us and the CDC in all its wisdom, hasn't even shut the barn door yet. They should have unilaterally ended all plane flights weeks ago but capitalism and commerce trump global public safety.
edit on 2027118pmThursdayf27Thu, 14 Aug 2014 23:27:20
-0500America/Chicago by signalfire because: addendum
a study in 2004 Multiple Ebola Virus Transmission Events and Rapid Decline of Central African Wildlife
Several human and animal Ebola outbreaks have occurred over the past 4 years in Gabon and the Republic of Congo. The human outbreaks consisted of multiple simultaneous epidemics caused by different viral strains, and each epidemic resulted from the handling of a distinct gorilla, chimpanzee, or duiker carcass. These animal populations declined markedly during human Ebola outbreaks, apparently as a result of Ebola infection. Recovered carcasses were infected by a variety of Ebola strains, suggesting that Ebola outbreaks in great apes result from multiple virus introductions from the natural host. Surveillance of animal mortality may help to predict and prevent human Ebola outbreaks.
Human Ebola virus (EBOV) infection causes hemorrhagic fever and death within a few days. The most lethal strains, causing up to 88% mortality, occur in Gabon, the Republic of Congo (RC), and the Democratic Republic of Congo (DRC) in central Africa, and belong to the Zaire subtype, which is one of four known EBOV subtypes. Together with Marburg virus, EBOV forms the Filoviridae family, a group of enveloped, nonsegmented, negative-strand RNA viruses .. Human Ebola outbreaks usually occur abruptly from an unidentified source, with subsequent spread from person to person (4). The first three known outbreaks of Ebola occurred between 1976 and 1979 in Zaire (now DRC) and Sudan, with 318, 284 and 34 cases, respectively. No further cases were recognized in Africa until late 1994. Since then, EBOV has appeared in human beings eight times, in several sub-Saharan African countries, including Côte d'Ivoire, DRC, Uganda, RC, and Gabon. Epidemiologic observations showed that chimpanzees were the source of one human case in Côte d'Ivoire in late 1994 and of an outbreak in Gabon in 1996. The recent high frequency of EBOV outbreaks in central Africa, associated with high lethality and serious social consequences, has made Ebola a major public health priority. Given the very rapid spread of the disease and the lack of a vaccine or effective therapy, the cornerstone of management remains the prevention and rapid control of outbreaks.
In the past two years, five human Ebola virus outbreaks of the Zaire subtype have occurred in western central Africa (Gabon and RC), with a total of 313 cases and 264 deaths. Epidemiologic investigations show that these outbreaks resulted from multiple introductions of the virus from different infected animal sources (Fig. 1A). The index cases (mainly hunters) were all infected when handling dead animals (gorilla, chimpanzee, or duiker), and subsequent transmission occurred by direct person-to-person contact, especially within families. We use the term “epidemic chain” to designate all cases of infection arising from a given index case and thus from a given animal carcass. At least ten epidemic chains were identified between October 2001 and May 2003.
To determine whether these outbreaks resulted from multiple introductions of a single viral strain or from separate introductions of several different viral strains, we sequenced the entire open reading frame of the EBOV glycoprotein (GP) gene in all available infected human samples. Nucleotide sequence variations were found among the viruses responsible for the different epidemic chains (Table 1) but not between samples from patients involved in a given epidemic chain. On the basis of genetic analysis, we identified eight viral strains, indicating that the five human outbreaks involved distinct animal sources and viral strains
We and local villagers noted a high number of animal carcasses in forested areas just before and during the 2001 human Ebola outbreaks in Gabon (Fig. 1B; table S1). Discovery of dead animals in the forest is normally rare, owing to rapid decomposition and the low number of nonpredatory deaths. Over a period of 8 months, we found or were informed by local villagers of at least 64 animal carcasses (gorillas, chimpanzees, and duikers) in the outbreak area of the Zadie region in Gabon (3000 km2), with a peak in November and December 2001; most were gorillas (36; table S1). Considering that we have observed that a gorilla carcass decomposes completely within one month in the tropical forest and that the carcasses were found in the vicinity of villages (within 2 hours walking distance), hundreds or even thousands of animals may have died in the thinly populated 3000 km2 of this forest region.
Gorilla (Gorilla gorilla) population censuses before and after the Kelle outbreak in 2003 in RC showed marked declines after the outbreak. Indices of the presence of gorilla and duiker (Cephalophus spp.)— sighting, hearing, and smelling the animals by trained trackers, and seeing dung, trails, or nests—fell by 50% between 2002 and 2003 in the 320-km2 Lossi sanctuary. Similarly, chimpanzee (Pan troglodytes) indices declined by 88%. Although a decline in indices does not indicate a similar decline in population densities, it is a robust indicator of a corresponding population decline. Corroborating this observation, eight groups of gorillas, 143 individuals in all, that had been monitored almost daily for 10 years in a 35-km2 region of the Lossi sanctuary, disappeared between October 2002 and January 2003. These animals have not been seen since, neither within the sanctuary nor elsewhere. Together, these data indicate temporal and spatial overlaps between human Ebola outbreaks and increased mortality among large wild mammals. Our data also confirm that Ebola outbreaks occur abruptly, exterminating exposed animal populations very rapidly and very locally (groups living in other parts of the sanctuary were barely affected). The duiker populations in the areas covered by our study may recover rapidly after an outbreak, owing to this species' rapid reproductive cycle. In contrast, the slow reproductive cycle of the great apes, together with hunting and poaching, may lead to their extinction in western central Africa. This last observation is supported by recent data indicating that ape populations in Gabon declined by more than half between 1983 and 2000. These findings are also consistent with those of a study conducted in the Taï Forest of Côte d'Ivoire, in which 11 of 43 members of a wild chimpanzee group disappeared in November 1994. One chimpanzee was positive by EBOV-specific immunohistochemical staining.
Using a combination of antigen detection, serology, DNA amplification, sequencing, immunohistochemical staining, and virus isolation, we confirmed that the gorillas, chimpanzees, and duiker carcasses were indeed infected by EBOV. In total, ten gorillas, three chimpanzees, and one duiker tested positive. Of interest, all muscle tissues were negative for EBOV-specific immunoglobulin G (IgG), indicating that the animals had died without developing specific IgG responses, similar to the situation in human infections in which a fatal outcome is characterized by impaired humoral responses and a failure to generate specific IgG.
Deaths in wildlife tended to precede and to be linked to human infections. For example, we detected EBOV in gorilla and chimpanzee carcasses in early December 2002 at the Lossi sanctuary, and the first human cases in the associated outbreak (Yembelengoye and Mvoula, 2003) appeared at the end of that month. In addition, serum from the index case (a survivor) in the last human epidemic in Mekambo (Grand Etoumbi, March 2002) was positive for EBOV-specific IgG, and EBOV virus sequences from the L gene were detected in bone-marrow samples of the gorilla identified as the source of this outbreak, conclusively linking the two cases.
Nucleotide sequence variations of the GP gene, distinct from those observed previously in humans, were also found in infected tissues from dead animals. We identified 11 different EBOV strains from human and animal samples between October 2001 and May 2003 in northern Gabon and in the region bordering RC. Molecular analyses based on estimated nucleotide substitution rates indicate that the four known EBOV subtypes diverged thousands of years ago and do not spread rapidly from one region to another of the central African forest block (16). A previous study focusing on the most divergent region of the GP gene shows that the EBOV isolates obtained from fatalities and survivors during the Kikwit outbreak in 1995 are genetically stable. Similarly, there were no nucleotide sequence variations in the GP and nucleoprotein genes among isolates from fatalities, survivors, and asymptomatically infected individuals during the 7-month Gabon outbreak in 1996–1997. The same study shows that the Booué sequence diverges from that of other Zaire subtype strains by only 1 to 2% (36 substitutions of a 2174-nucleotide fragment), despite the 20-year interval and 3000-km distance between the Booué 1996 and Zaire 1976/1995 outbreaks. The very high genetic stability of EBOV, and our findings of many different EBOV sequences that do not vary during transmission along a given human epidemic chain indicate that multiple independent introductions of the virus from its natural host to great apes had occurred.
Humans and duikers scavenging for meat probably become infected by contact with dead apes. However, we found that animal carcasses left in the forest are not infectious after 3 to 4 days. Therefore, virus transmission between different groups of apes is unlikely, because infectivity is short-lived and physical contact between individuals from different groups is rare. Different strains of EBOV may be widespread throughout the forests of central Africa, with simultaneous infection of great apes occurring from still unknown natural hosts under particular but unknown environmental conditions. Ebola outbreaks in great apes have always been reported at the beginning of the dry seasons (December 1995 in Mayibout, July 1996 in Booué, July 2001 in Mekambo, December 2001 in Kelle, and December 2002 in the second Kelle outbreak). Thus, Ebola outbreaks probably do not occur as a single outbreak spreading throughout the Congo basin as others have proposed, but are due to multiple episodic infection of great apes from the reservoir.
The simultaneous occurrence of multiple epidemic chains in animals complicates the work of outbreak-response teams, because new cases may appear in unmonitored populations. Almost all human Ebola outbreaks in Gabon and RC have been linked to the handling of dead animals by villagers or hunters, and increased animal mortality always preceded the first human cases. A monitoring network for large-animal mortality rates is needed for rapid implementation of EBOV prevention measures when increased deaths are observed among wild animals living near human communities.
Several human and animal Ebola outbreaks have occurred over the past 4 years in Gabon and the Republic of Congo. The human outbreaks consisted of multiple simultaneous epidemics caused by different viral strains, and each epidemic resulted from the handling of a distinct gorilla, chimpanzee, or duiker carcass. These animal populations declined markedly during human Ebola outbreaks, apparently as a result of Ebola infection. Recovered carcasses were infected by a variety of Ebola strains, suggesting that Ebola outbreaks in great apes result from multiple virus introductions from the natural host. Surveillance of animal mortality may help to predict and prevent human Ebola outbreaks.
Human Ebola virus (EBOV) infection causes hemorrhagic fever and death within a few days. The most lethal strains, causing up to 88% mortality, occur in Gabon, the Republic of Congo (RC), and the Democratic Republic of Congo (DRC) in central Africa, and belong to the Zaire subtype, which is one of four known EBOV subtypes. Together with Marburg virus, EBOV forms the Filoviridae family, a group of enveloped, nonsegmented, negative-strand RNA viruses .. Human Ebola outbreaks usually occur abruptly from an unidentified source, with subsequent spread from person to person (4). The first three known outbreaks of Ebola occurred between 1976 and 1979 in Zaire (now DRC) and Sudan, with 318, 284 and 34 cases, respectively. No further cases were recognized in Africa until late 1994. Since then, EBOV has appeared in human beings eight times, in several sub-Saharan African countries, including Côte d'Ivoire, DRC, Uganda, RC, and Gabon. Epidemiologic observations showed that chimpanzees were the source of one human case in Côte d'Ivoire in late 1994 and of an outbreak in Gabon in 1996. The recent high frequency of EBOV outbreaks in central Africa, associated with high lethality and serious social consequences, has made Ebola a major public health priority. Given the very rapid spread of the disease and the lack of a vaccine or effective therapy, the cornerstone of management remains the prevention and rapid control of outbreaks.
In the past two years, five human Ebola virus outbreaks of the Zaire subtype have occurred in western central Africa (Gabon and RC), with a total of 313 cases and 264 deaths. Epidemiologic investigations show that these outbreaks resulted from multiple introductions of the virus from different infected animal sources (Fig. 1A). The index cases (mainly hunters) were all infected when handling dead animals (gorilla, chimpanzee, or duiker), and subsequent transmission occurred by direct person-to-person contact, especially within families. We use the term “epidemic chain” to designate all cases of infection arising from a given index case and thus from a given animal carcass. At least ten epidemic chains were identified between October 2001 and May 2003.
To determine whether these outbreaks resulted from multiple introductions of a single viral strain or from separate introductions of several different viral strains, we sequenced the entire open reading frame of the EBOV glycoprotein (GP) gene in all available infected human samples. Nucleotide sequence variations were found among the viruses responsible for the different epidemic chains (Table 1) but not between samples from patients involved in a given epidemic chain. On the basis of genetic analysis, we identified eight viral strains, indicating that the five human outbreaks involved distinct animal sources and viral strains
We and local villagers noted a high number of animal carcasses in forested areas just before and during the 2001 human Ebola outbreaks in Gabon (Fig. 1B; table S1). Discovery of dead animals in the forest is normally rare, owing to rapid decomposition and the low number of nonpredatory deaths. Over a period of 8 months, we found or were informed by local villagers of at least 64 animal carcasses (gorillas, chimpanzees, and duikers) in the outbreak area of the Zadie region in Gabon (3000 km2), with a peak in November and December 2001; most were gorillas (36; table S1). Considering that we have observed that a gorilla carcass decomposes completely within one month in the tropical forest and that the carcasses were found in the vicinity of villages (within 2 hours walking distance), hundreds or even thousands of animals may have died in the thinly populated 3000 km2 of this forest region.
Gorilla (Gorilla gorilla) population censuses before and after the Kelle outbreak in 2003 in RC showed marked declines after the outbreak. Indices of the presence of gorilla and duiker (Cephalophus spp.)— sighting, hearing, and smelling the animals by trained trackers, and seeing dung, trails, or nests—fell by 50% between 2002 and 2003 in the 320-km2 Lossi sanctuary. Similarly, chimpanzee (Pan troglodytes) indices declined by 88%. Although a decline in indices does not indicate a similar decline in population densities, it is a robust indicator of a corresponding population decline. Corroborating this observation, eight groups of gorillas, 143 individuals in all, that had been monitored almost daily for 10 years in a 35-km2 region of the Lossi sanctuary, disappeared between October 2002 and January 2003. These animals have not been seen since, neither within the sanctuary nor elsewhere. Together, these data indicate temporal and spatial overlaps between human Ebola outbreaks and increased mortality among large wild mammals. Our data also confirm that Ebola outbreaks occur abruptly, exterminating exposed animal populations very rapidly and very locally (groups living in other parts of the sanctuary were barely affected). The duiker populations in the areas covered by our study may recover rapidly after an outbreak, owing to this species' rapid reproductive cycle. In contrast, the slow reproductive cycle of the great apes, together with hunting and poaching, may lead to their extinction in western central Africa. This last observation is supported by recent data indicating that ape populations in Gabon declined by more than half between 1983 and 2000. These findings are also consistent with those of a study conducted in the Taï Forest of Côte d'Ivoire, in which 11 of 43 members of a wild chimpanzee group disappeared in November 1994. One chimpanzee was positive by EBOV-specific immunohistochemical staining.
Using a combination of antigen detection, serology, DNA amplification, sequencing, immunohistochemical staining, and virus isolation, we confirmed that the gorillas, chimpanzees, and duiker carcasses were indeed infected by EBOV. In total, ten gorillas, three chimpanzees, and one duiker tested positive. Of interest, all muscle tissues were negative for EBOV-specific immunoglobulin G (IgG), indicating that the animals had died without developing specific IgG responses, similar to the situation in human infections in which a fatal outcome is characterized by impaired humoral responses and a failure to generate specific IgG.
Deaths in wildlife tended to precede and to be linked to human infections. For example, we detected EBOV in gorilla and chimpanzee carcasses in early December 2002 at the Lossi sanctuary, and the first human cases in the associated outbreak (Yembelengoye and Mvoula, 2003) appeared at the end of that month. In addition, serum from the index case (a survivor) in the last human epidemic in Mekambo (Grand Etoumbi, March 2002) was positive for EBOV-specific IgG, and EBOV virus sequences from the L gene were detected in bone-marrow samples of the gorilla identified as the source of this outbreak, conclusively linking the two cases.
Nucleotide sequence variations of the GP gene, distinct from those observed previously in humans, were also found in infected tissues from dead animals. We identified 11 different EBOV strains from human and animal samples between October 2001 and May 2003 in northern Gabon and in the region bordering RC. Molecular analyses based on estimated nucleotide substitution rates indicate that the four known EBOV subtypes diverged thousands of years ago and do not spread rapidly from one region to another of the central African forest block (16). A previous study focusing on the most divergent region of the GP gene shows that the EBOV isolates obtained from fatalities and survivors during the Kikwit outbreak in 1995 are genetically stable. Similarly, there were no nucleotide sequence variations in the GP and nucleoprotein genes among isolates from fatalities, survivors, and asymptomatically infected individuals during the 7-month Gabon outbreak in 1996–1997. The same study shows that the Booué sequence diverges from that of other Zaire subtype strains by only 1 to 2% (36 substitutions of a 2174-nucleotide fragment), despite the 20-year interval and 3000-km distance between the Booué 1996 and Zaire 1976/1995 outbreaks. The very high genetic stability of EBOV, and our findings of many different EBOV sequences that do not vary during transmission along a given human epidemic chain indicate that multiple independent introductions of the virus from its natural host to great apes had occurred.
Humans and duikers scavenging for meat probably become infected by contact with dead apes. However, we found that animal carcasses left in the forest are not infectious after 3 to 4 days. Therefore, virus transmission between different groups of apes is unlikely, because infectivity is short-lived and physical contact between individuals from different groups is rare. Different strains of EBOV may be widespread throughout the forests of central Africa, with simultaneous infection of great apes occurring from still unknown natural hosts under particular but unknown environmental conditions. Ebola outbreaks in great apes have always been reported at the beginning of the dry seasons (December 1995 in Mayibout, July 1996 in Booué, July 2001 in Mekambo, December 2001 in Kelle, and December 2002 in the second Kelle outbreak). Thus, Ebola outbreaks probably do not occur as a single outbreak spreading throughout the Congo basin as others have proposed, but are due to multiple episodic infection of great apes from the reservoir.
The simultaneous occurrence of multiple epidemic chains in animals complicates the work of outbreak-response teams, because new cases may appear in unmonitored populations. Almost all human Ebola outbreaks in Gabon and RC have been linked to the handling of dead animals by villagers or hunters, and increased animal mortality always preceded the first human cases. A monitoring network for large-animal mortality rates is needed for rapid implementation of EBOV prevention measures when increased deaths are observed among wild animals living near human communities.
www.wnd.com...
By Jane M. Orient, M.D., director of the Association of American Physicians and Surgeons (ron paul is a member)
claims that
there is evidence that airborne transmission can occur in some circumstances, as noted in a Pathogen Safety Data Sheet from the Public Health Agency of Canada
and says
Whatever the source, epidemics like Ebola must not be viewed with complacency.
By Jane M. Orient, M.D., director of the Association of American Physicians and Surgeons (ron paul is a member)
claims that
there is evidence that airborne transmission can occur in some circumstances, as noted in a Pathogen Safety Data Sheet from the Public Health Agency of Canada
and says
Whatever the source, epidemics like Ebola must not be viewed with complacency.
a reply to: Darkblade71
What...with potentially hundreds of millions of doses of this new 'super-duper-experimental-ebola-drug' they could see flying off the shelves like there's no tomorrow?
I hope you're right, and i'm wrong...but don't count on it.
I think it will stay contained for the most part in Africa.
What...with potentially hundreds of millions of doses of this new 'super-duper-experimental-ebola-drug' they could see flying off the shelves like there's no tomorrow?
I hope you're right, and i'm wrong...but don't count on it.
www.cdc.gov...
in cdc website above low risk exposure were found but also people with no known exposure were found also found ??!! in the current outbreak in west africa (very serious)
johngaltfla.com... dividual/
quote cdc update in aug 7 based on aug 4 data:
High risk exposures
A high risk exposure includes any of the following:
•Percutaneous, e.g. the needle stick, or mucous membrane exposure to body fluids of EVD patient
•Direct care or exposure to body fluids of an EVD patient without appropriate personal protective equipment (PPE)
•Laboratory worker processing body fluids of confirmed EVD patients without appropriate PPE or standard biosafety precautions
•Participation in funeral rites which include direct exposure to human remains in the geographic area where outbreak is occurring without appropriate PPE
Low risk exposures
A low risk exposure includes any of the following
•Household member or other casual contact1 with an EVD patient
•Providing patient care or casual contact1 without high-risk exposure with EVD patients in health care facilities in EVD outbreak affected countries*
No known exposure
Persons with no known exposure were present in an EVD outbreak affected country* in the past 21 days with no low risk or high risk exposures.
1 Casual contact is defined as a) being within approximately 3 feet (1 meter) or within the room or care area for a prolonged period of time (e.g., healthcare personnel, household members) while not wearing recommended personal protective equipment (i.e., droplet and contact precautions–see Infection Prevention and Control Recommendations); or b) having direct brief contact (e.g., shaking hands) with an EVD case while not wearing recommended perso
nal protective equipment (i.e., droplet and contact precautions–see Infection Prevention and Control Recommendations). At this time, brief interactions, such as walking by a person or moving through a hospital, do not constitute casual contact.
* Outbreak affected countries include Guinea, Liberia, Sierra Leone, and Lagos, Nigeria, as of 4-August-2014
"
!?
in cdc website above low risk exposure were found but also people with no known exposure were found also found ??!! in the current outbreak in west africa (very serious)
johngaltfla.com... dividual/
edit on 15-8-2014 by Starbucks because: (no reason given)
edit on 15-8-2014 by Starbucks because: (no reason
given)
quote cdc update in aug 7 based on aug 4 data:
High risk exposures
A high risk exposure includes any of the following:
•Percutaneous, e.g. the needle stick, or mucous membrane exposure to body fluids of EVD patient
•Direct care or exposure to body fluids of an EVD patient without appropriate personal protective equipment (PPE)
•Laboratory worker processing body fluids of confirmed EVD patients without appropriate PPE or standard biosafety precautions
•Participation in funeral rites which include direct exposure to human remains in the geographic area where outbreak is occurring without appropriate PPE
Low risk exposures
A low risk exposure includes any of the following
•Household member or other casual contact1 with an EVD patient
•Providing patient care or casual contact1 without high-risk exposure with EVD patients in health care facilities in EVD outbreak affected countries*
No known exposure
Persons with no known exposure were present in an EVD outbreak affected country* in the past 21 days with no low risk or high risk exposures.
1 Casual contact is defined as a) being within approximately 3 feet (1 meter) or within the room or care area for a prolonged period of time (e.g., healthcare personnel, household members) while not wearing recommended personal protective equipment (i.e., droplet and contact precautions–see Infection Prevention and Control Recommendations); or b) having direct brief contact (e.g., shaking hands) with an EVD case while not wearing recommended perso
nal protective equipment (i.e., droplet and contact precautions–see Infection Prevention and Control Recommendations). At this time, brief interactions, such as walking by a person or moving through a hospital, do not constitute casual contact.
* Outbreak affected countries include Guinea, Liberia, Sierra Leone, and Lagos, Nigeria, as of 4-August-2014
"
!?
edit on 15-8-2014 by Starbucks because: (no reason given)
From WHO
Well I can say that I would not want to be on one of these flights that have now been given the 'Green Light' to fly would anyone else especially on the off change you are in the adjoining seat? I think people will make their own decision on this and how confident they are with this announcement.
WHO: Air travel is low-risk for Ebola transmission
Note for the media
14 AUGUST 2014 ¦ GENEVA - The World Health Organization (WHO) today reiterated its position that the risk of transmission of Ebola virus disease during air travel remains low.
“Unlike infections such as influenza or tuberculosis, Ebola is not airborne,” says Dr Isabelle Nuttall, Director of WHO Global Capacity Alert and Response. “It can only be transmitted by direct contact with the body fluids of a person who is sick with the disease.”
On the small chance that someone on the plane is sick with Ebola, the likelihood of other passengers and crew having contact with their body fluids is even smaller. Usually when someone is sick with Ebola, they are so unwell that they cannot travel. WHO is therefore advising against travel bans to and from affected countries.
Well I can say that I would not want to be on one of these flights that have now been given the 'Green Light' to fly would anyone else especially on the off change you are in the adjoining seat? I think people will make their own decision on this and how confident they are with this announcement.
dr jane orient says airborne did happen. she says that cdc director claimed in aug 1 that no possibility of ebola in us but on aug 7 he says it could
big time betwweb aug 1 and 7 they flew brently to us.
the cdc knew in their update dated aug 7 that in 21 previous days transmittion with no contact happened (so they knew in july 15 )but his statement in aug 1.
casual contact in cdc margin of page is 3 feet away from patient, while unknown exposure is far away
the cdc knew in their update dated aug 7 that in 21 previous days transmittion with no contact happened (so they knew in july 15 )but his statement in aug 1.
casual contact in cdc margin of page is 3 feet away from patient, while unknown exposure is far away
I've updated the Ebola outbreak charts, including the projection charts, with the new data WHO released today. That data covers through August 13,
2014. Click any graphic below to see it larger.
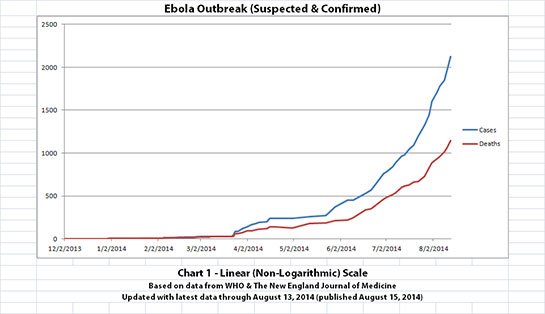
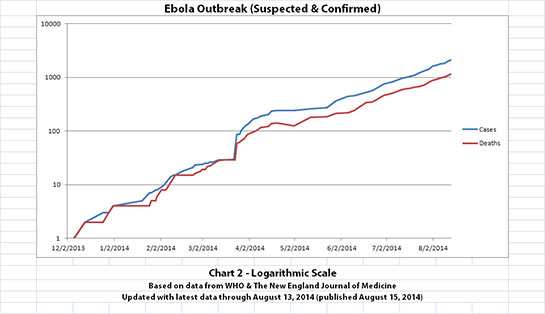
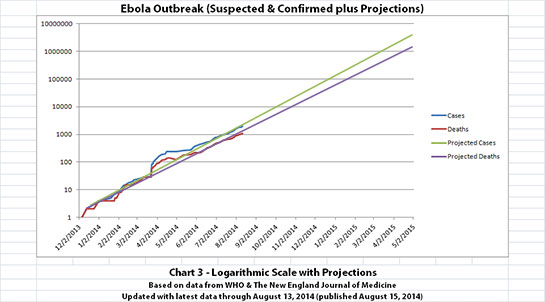
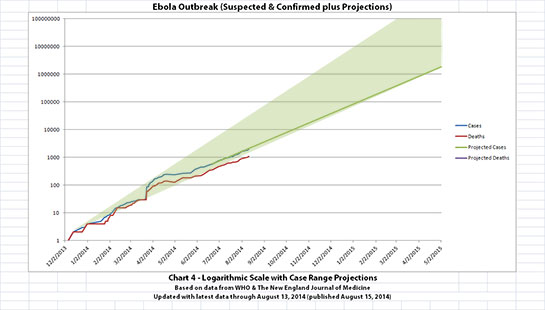
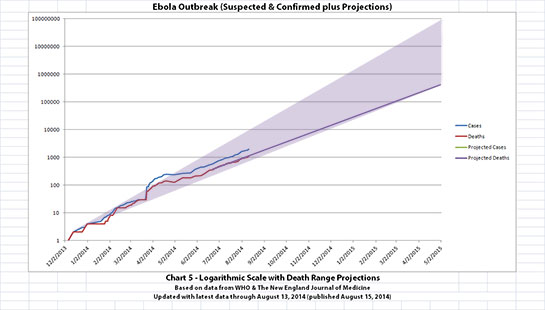
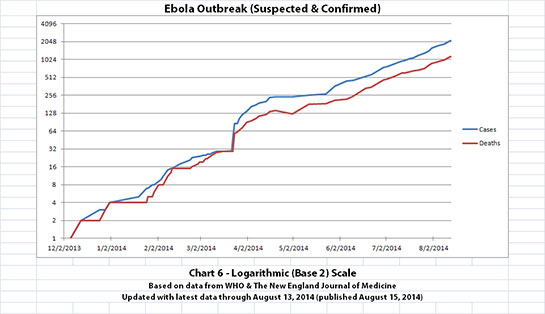
The charts and future projections were done by me, not WHO. I am not an Ebola expert, epidemiologist, virologist, or MD, and I hope the projections are wrong. Charts 1, 2, and 6 show cases and deaths officially reported. The y-axis is linear in Chart 1. In Chart 2, it is in powers of 10. In Chart 6 it is in powers of 2. I updated known data in Charts 3-5 but left the projections the same as on August 8, 2014.
The projections are controversial. Most appreciate them, but a few think they grossly under or overestimate or try to project what cannot be projected. Please do not do anything you might regret based on these projections. Efforts to contain, quarantine, treat, prevent, or cure Ebola will hopefully prove them wrong.
Data through August 13, 2014 came from news updates on the following WHO and The New England Journal of Medicine websites:
SOURCE: WHO website 1
SOURCE: WHO website 2
SOURCE: The New England Journal of Medicine
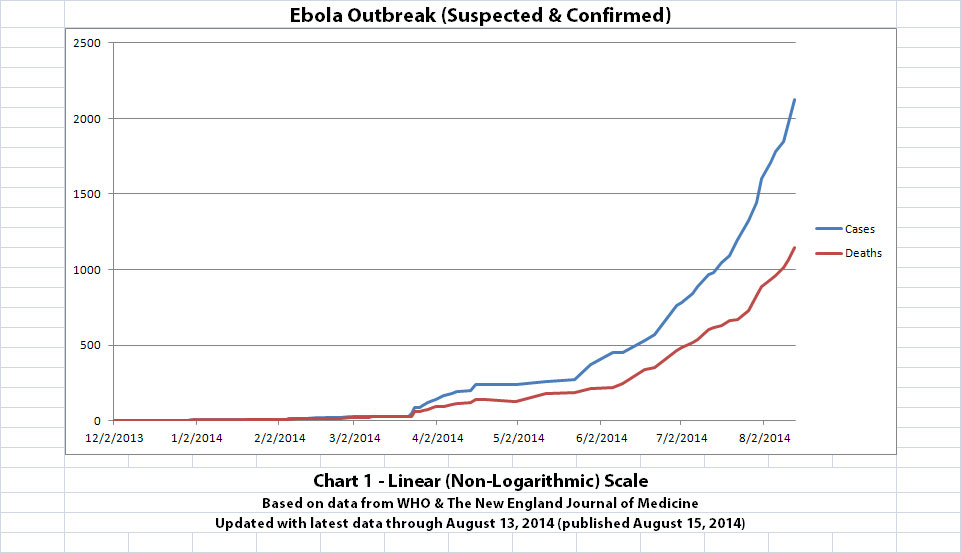
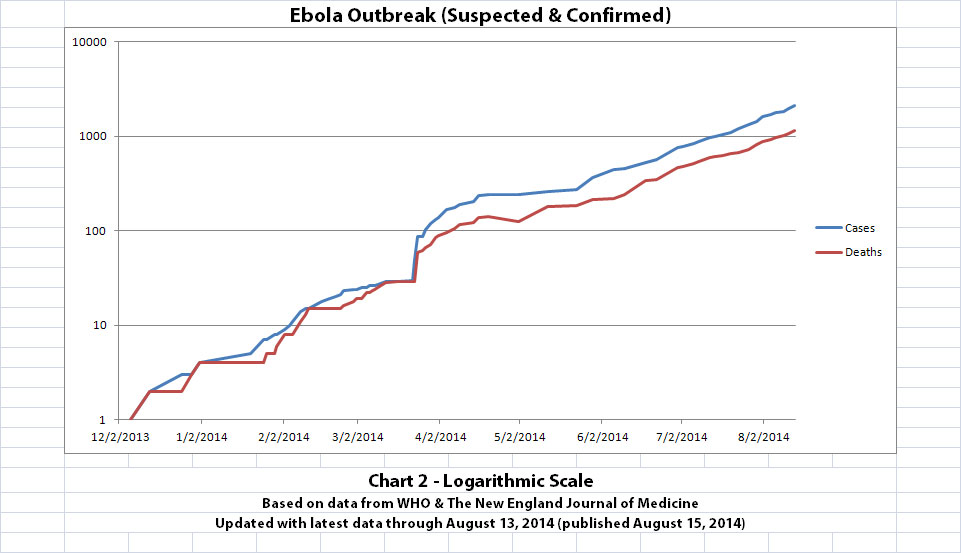
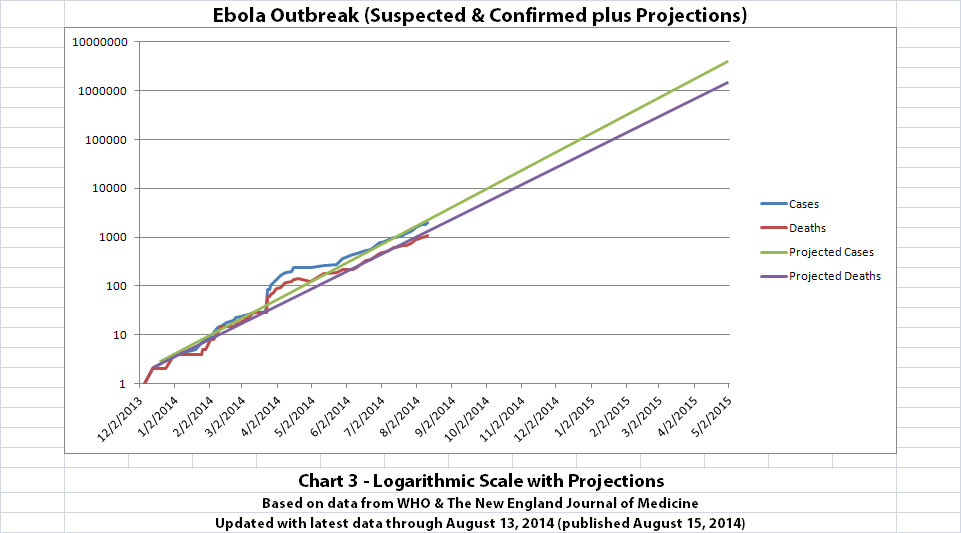
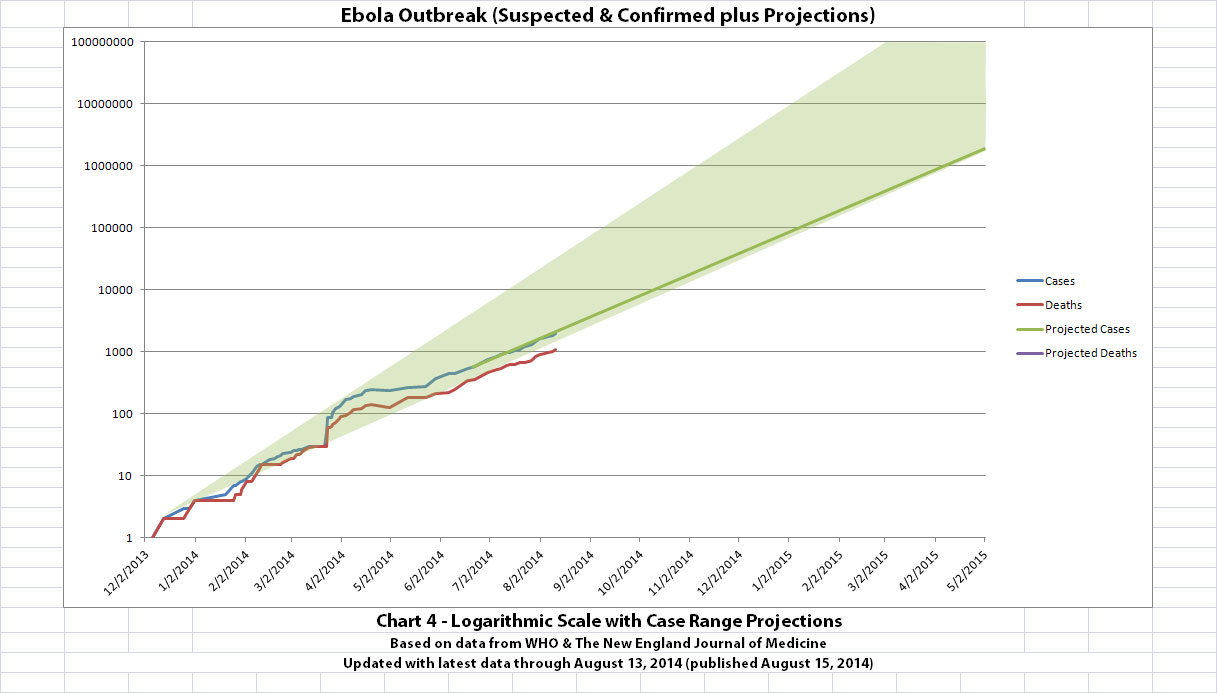
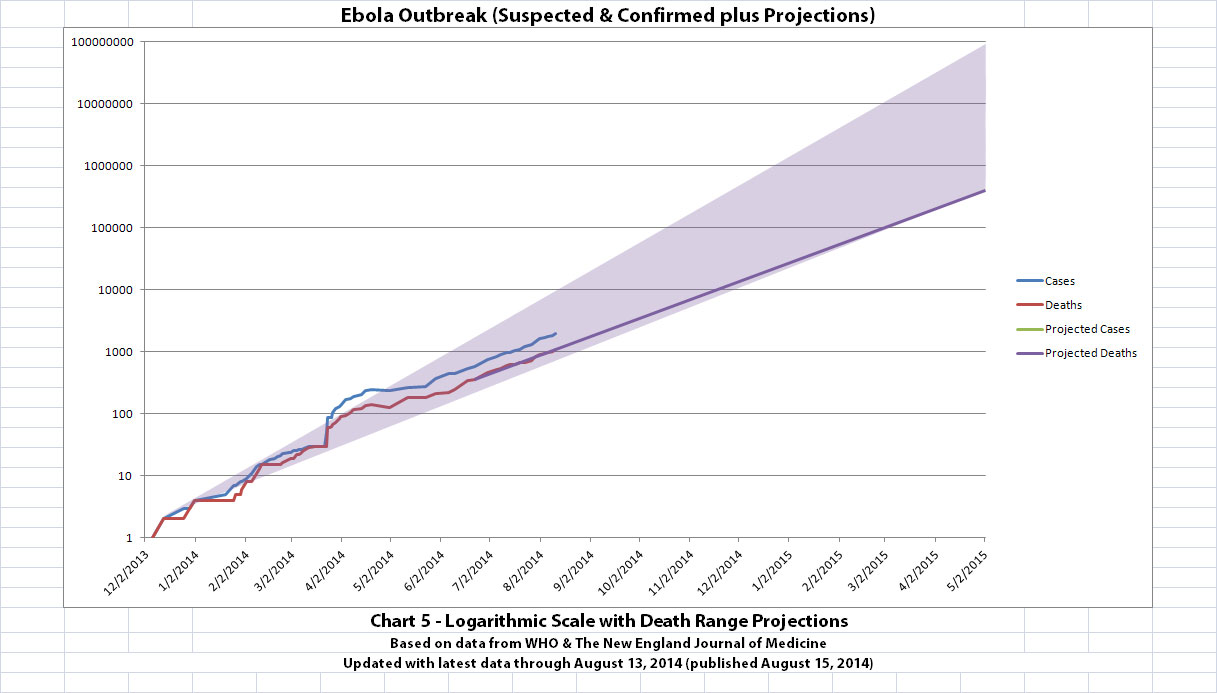
The charts and future projections were done by me, not WHO. I am not an Ebola expert, epidemiologist, virologist, or MD, and I hope the projections are wrong. Charts 1, 2, and 6 show cases and deaths officially reported. The y-axis is linear in Chart 1. In Chart 2, it is in powers of 10. In Chart 6 it is in powers of 2. I updated known data in Charts 3-5 but left the projections the same as on August 8, 2014.
The projections are controversial. Most appreciate them, but a few think they grossly under or overestimate or try to project what cannot be projected. Please do not do anything you might regret based on these projections. Efforts to contain, quarantine, treat, prevent, or cure Ebola will hopefully prove them wrong.
Data through August 13, 2014 came from news updates on the following WHO and The New England Journal of Medicine websites:
SOURCE: WHO website 1
SOURCE: WHO website 2
SOURCE: The New England Journal of Medicine
new topics
-
Holy Cow! Erm...Six Legged Turkey!!
World Sports: 3 hours ago -
Ben Habib has Left Reform UK
Regional Politics: 6 hours ago -
Turkey Day Rhyme…
Short Stories: 6 hours ago -
Can someone please translate Biden's speech?
US Political Madness: 7 hours ago -
NIH Chief Confesses COVID Initiatives Were "Completely Made Up " OMG Investigates
Health & Wellness: 7 hours ago -
Awesome Dip Recipe
Food and Cooking: 10 hours ago -
Vladimir Putin's speech at the meeting of the CSTO Collective Security Council
World War Three: 11 hours ago
top topics
-
NIH Chief Confesses COVID Initiatives Were "Completely Made Up " OMG Investigates
Health & Wellness: 7 hours ago, 9 flags -
Vladimir Putin's speech at the meeting of the CSTO Collective Security Council
World War Three: 11 hours ago, 8 flags -
Traveling the world with no passport
Social Issues and Civil Unrest: 15 hours ago, 7 flags -
Can someone please translate Biden's speech?
US Political Madness: 7 hours ago, 7 flags -
Awesome Dip Recipe
Food and Cooking: 10 hours ago, 5 flags -
Turkey Day Rhyme…
Short Stories: 6 hours ago, 5 flags -
Ben Habib has Left Reform UK
Regional Politics: 6 hours ago, 4 flags -
Holy Cow! Erm...Six Legged Turkey!!
World Sports: 3 hours ago, 2 flags
active topics
-
The Party of Peace - Trump Cabinet Picks Targeted with Death Threats
US Political Madness • 37 • : chr0naut -
V.P. Kamala Harris releases a video and nobody understands why
US Political Madness • 85 • : chr0naut -
Traveling the world with no passport
Social Issues and Civil Unrest • 9 • : KnowItAllKnowNothin -
NIH Chief Confesses COVID Initiatives Were "Completely Made Up " OMG Investigates
Health & Wellness • 14 • : KnowItAllKnowNothin -
Vladimir Putin's speech at the meeting of the CSTO Collective Security Council
World War Three • 59 • : lostgirl -
Holy Cow! Erm...Six Legged Turkey!!
World Sports • 2 • : RazorV66 -
Awesome Dip Recipe
Food and Cooking • 5 • : JJproductions -
Turkey Day Rhyme…
Short Stories • 4 • : JJproductions -
Mood Music Part VI
Music • 3721 • : BrucellaOrchitis -
Interesting Video-UFO?
Aliens and UFOs • 20 • : BrucellaOrchitis